Title:Demography and its effects on genomic variation in crop domestication
背景 :
超过两千种植物被驯化;也就是说,它们通过人类培养在形态和遗传上进行了修饰。与驯化相关的修饰包括一系列形态变化,例如增强的稳健性,产生较少但较大的果实(或谷粒),以及改变的种子休眠和扩散等。
然而,形态变化并不会在驯化后停止,因为初期作物通常会分散到新的位置,从而产生新的生物和非生物胁迫。这些压力通过额外的表型和遗传变化推动了局部适应。
文章结构:
该综述主要从三方面阐述人类对作物驯化基因组变异的影响。第一方面,驯化的阶段与持续时间的角度,第二方面,人类塑造了不利变体的范围,第三人类导致的一些不确定结构变异对驯化的影响。
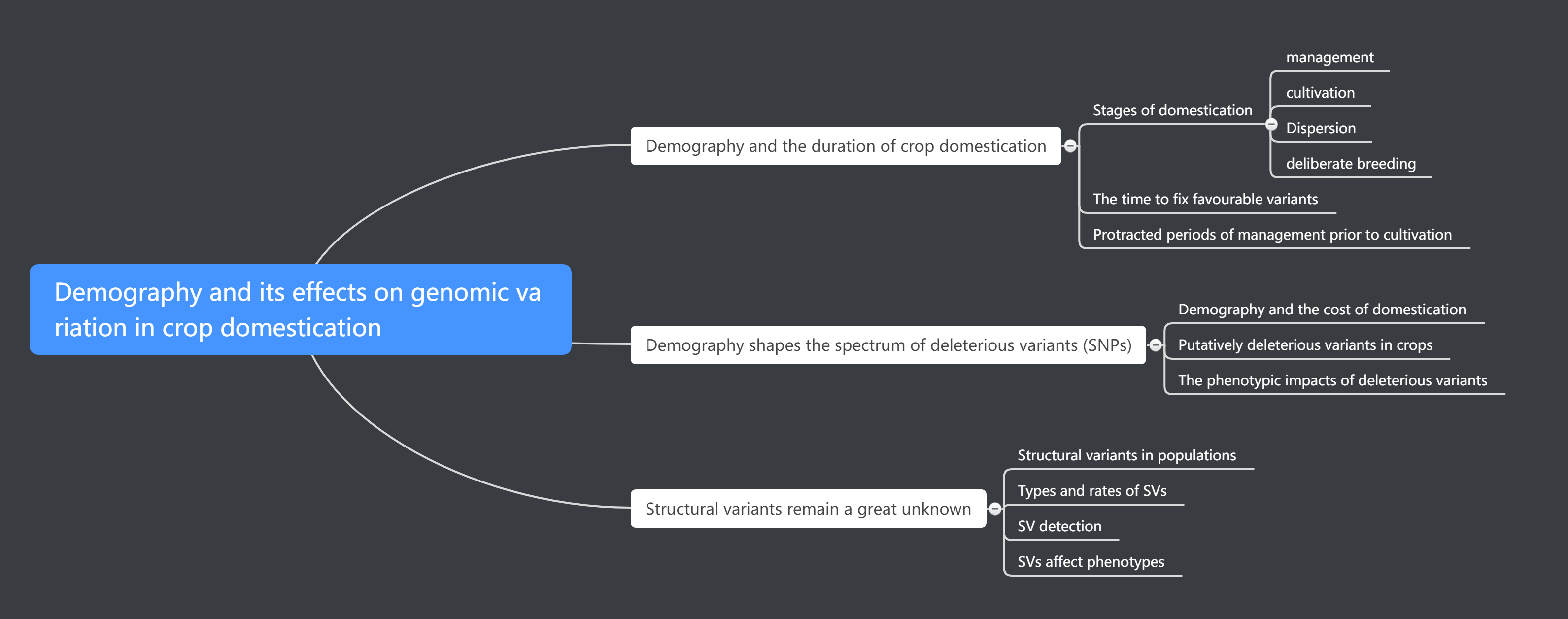 Demography and its effect.png-271.9kB
Demography and its effect.png-271.9kB
人类影响驯化的阶段与持续时间
驯化的阶段
 此处输入图片的描述
此处输入图片的描述
主要可以分为4个阶段,阶段一管理,在正式种植之前,人类管理和收获野生植物种群的可能性。第二阶段,驯化耕种,通过驯化瓶颈扼杀遗传多样性并同时增加驯化等位基因频率的过程。第三阶段,分离,驯化的地理扩张,往往需要适应新的环境。第四阶段,育种过去几百年来发生的一个过程。植物在这些阶段的持续时间或进行的程度方面是相同的。例如,大多数谷类作物经历了强烈的原位选择和显着的遗传瓶颈(第2阶段),但许多多年生作物没有。
读到这里时候,这个简单的阶段图很吸引我,简单明了了表达了很多信息。但万万没想到的是,后面两个大的方面作者也一直紧紧扣住这个阶段图,将下面要表达的内容与其相结合。越读到下文,越能体会到该图的切到好处的地方。
产生有利变异的时间
当今的研究主要是关注于阶段2驯化的时间。然后目前比较流行的方法有两种:种群模型分析(using SNP data)和化石分析。
然后文章很详细的介绍并简单的比较了两种方法优与劣势。简而言之,有两个观点可以从中得出:1)不是一成不变的随着时间的推移可能有助于解决基于化石和建模的固定时间推断之间的明显差异 2)两种方法可能反映了等位基因和表型之间的不完全相关性。
驯化培养前的长期管理期
主要讲述,使用马尔可夫聚结方法(或“SMC”方法)来推断人对于驯化前管理时期的影响,然后阐明了很多该方法的好处与坏处。作者总结出,改研究方向的后续步骤将包括进一步应用基于SMC的方法来处理来自更多作物物种的基因组数据,模拟研究测试SMC方法在合理的演示场景下的推理稳健性,以及人口模拟评估经验数据是否适合基于SMC的人口下降的推论。(这部分比较理论)
往往reviewer很喜欢你从两面去思考一个问题,他们喜欢看你怎么从辩证的角度从好和坏去解答该问题。
人类影响有害变体的范围
人类影响与驯化成本
Ne: 有效群体规模是“为下一代贡献后代的人口中的个体数量”。
“驯化成本”是指品种内的遗传负荷增加,理论上可以通过鉴定作物基因组中的有害变异来研究。为了更好地了解群体规模的减少情况,有助于有害等位基因的积累,首先考虑野生祖先群体中突变的适应效应(DFE)的分布是很有帮助的。突变的影响范围从强烈有害到高度有利。大多数作物野生祖先的突变具有从零(即选择性中性)到轻微有害的适应性影响。通常在具有大Ne的群体中可以发现低频率发现轻微有害突变,因为选择是在大量群体中有明显的效应。
首先,瓶颈效应减少了遗传变异。通过减少遗传变异,瓶颈效应可能会去除许多(也许是大多数)略有缺失的遗传变异。其次,选择效果会变得不那么有效,所以由于遗传漂变,未丢失的有害变体往往会升高到更高的频率。
在作物中推定有害的变体
有几种方法可以执行基于WGS的有害变异的预测;他们中的大多数认为缺失的变异是进化上保守的基因组区域内的核苷酸变化。基于这些方法的应用实例,与其野生近缘种相比,大多数作物都经过检验出含有丰富比例的有害SNP变种。木薯(Manihotesculenta)是目前有害基因的冠军,因为单个木薯个体平均比相应的野生木薯基因具有大约26%的预测有害等位基因。由于不同的因素,有害突变的积累因作物而异。一个是人口历史,持续的Ne下降(第1阶段)加剧有害变体的积累。积累也将根据驯化瓶颈的强度和持续时间而变化。这种积累也受到DFE的影响物种之间以及野生和栽培形式之间可能不同。另一个因素是交配系统,因为它影响有害变体的可能保留或清除。例如,克隆繁殖可以隐藏杂合状态下的隐性有害变体,但自体受精不能。鉴于这些考虑因素,木薯中有害变异显着增加26%可能是由于其克隆繁殖以及驯化期间严重的种群瓶颈。
有害变体的表型影响
“驯化成本”不仅仅是人口历史的成果,而且是改善作物通过育种去除有害变种的潜在途径。但一个很重要的问题是,如何决定成千上万的有害变种中哪一种是作物改良的适当目标。有趣的是,作物可能能够逃脱他们的一些有害变体,当作物从其驯化中心分散到新的地理区域时,这是一种特别重要的进化力量。分散这个阶段需要适应新的环境,这可以通过在作物内进行选择或通过从当地适应的野生种群中进行等位基因的渐渗来实现。当Ne中两个群体不同时,基因渐渗可以改变遗传负荷,因此改变在他们的有害变异的遗传的负荷。
结构变异仍然是一个未知数
与驯化相关的人类影响也决定了作物中SV的数量和频率。然而,目前关于作物或其野生近缘种中SV的种群和进化动态的信息很少。
群体中的结构变异
大多数SV基因座失去遗传变异,不再在作物内分离。随着作物的扩大,选择将针对这些高频率变异,突变将引入新的SV。该模型预测比它们的野生亲缘,作物将具有较少的多态性在第1阶段和第2阶段之后,SV位点,并且剩余的衍生和新引入的SV将有助于贡献与驯化的成本。
随着三代技术的发展,无论是人类的疾病还是植物的性状研究,SVs变异这一部分都会成为以后的亮点。
SV的类型和速率
SV的主要类型包括:insertions, deletions, duplications, inversions, trans- locations and complex events,TEs,CNVs。
SVs提供有关植物驯化历史和表型变异的遗传贡献的信息的潜力是巨大的。
SVs 探测
通过基于测序深度的short-read WGS data来探测,未来long-reads sequence将会有很多优势。还有相应的泛基因组的应用去检测完整的SVs。
SVs影响表型
大多数作物适应是由SV驱动的。如果这是真的,大多数关于作物适应和驯化的研究都忽略了关键的基因组变异的影响。
小结
该文章思路清晰,从一个我们很常见的角度,人类对作物驯化的影响来进行一系列的阐述。个人对最后SVs和驯化遗传的相关联系是非常感兴趣的,因为目前这方面的研究还是比较欠缺,上升提高的空间还很多。









网友评论